近日,Plant Biotechnology Journal在线发表了我室胡春根教授和张金智教授课题组的重要研究成果。该研究首次深入解析了 miR156-SPLs 基因模块如何通过调控赤霉素(GA)生物合成,协同控制柑橘开花与株高发育的双重机制。这一发现为攻克柑橘漫长童期和树体高大这两大制约产业发展的关键瓶颈,提供了全新的分子靶标和理论依据,对加速柑橘矮化密植新品种培育、提升柑橘生产效益具有重大意义。

开花标志着植物成功从营养生长转向生殖生长,是决定繁殖效率的核心环节。然而,多年生木本柑橘普遍存在漫长的童期,严重阻碍了良种选育与推广进程。同时,传统柑橘树体高大、树冠郁闭,显著增加了修剪、喷药、采收等环节的管理难度和成本。相比之下,矮化树型可实现更高密度种植、提升光合效率、方便机械化作业、降低生产成本并提高果实品质与产量。因此,发掘能同时促进早花(缩短童期)和诱导矮化的关键调控因子,是提升柑橘产业效率和效益的核心需求。
该研究中,研究人员发现过表达柑橘 miR156 家族成员 Ci-miR156c,延迟开花(有助于缩短童期研究)并显著降低株高(诱导矮化),展现了该模块调控关键农艺性状的巨大潜力。研究进一步揭示了其作用机制:Ci-miR156c 通过抑制不同的 SPL 转录因子(CiSPLs),分别调控开花与株高通路。在调控开花通路中, Ci-miR156c-CiSPL7 模块靶向激活开花关键因子 CiFD,从而控制柑橘成花转变。Ci-miR156c 通过 CiSPL6 和 CiSPL3 两条路径抑制株高:首先直接靶向抑制赤霉素生物合成的关键基因 CiGA20ox2,降低活性 GA 水平;其次通过 CiSPL3 调控 KNOX 家族基因 CiKN6,进而影响 CiGA20ox2 的表达。尤为重要的是,过表达 CiKN6 本身即可直接导致柑橘植株矮化,有力证实了其在株高控制中的核心作用。外源施用赤霉素(GA)及其生物合成抑制剂的实验结果,进一步证实了 miR156-SPLs 模块通过抑制 GA 合成途径实现对株高的控制。具体研究结果如下:
1. Ci-miR156c参与了调控开花和株高
表型观察显示,与对照组相比,Ci-miR156c 超量表达植株表现出植株矮化、刺长缩短和叶面积减小(图1a-d)。对株高、叶片数量、平均节间长、刺长和叶面积的统计分析进一步证实,超量表达系的株高、平均节间长、刺长和叶面积均显著低于对照组,而叶片数量无显著差异(图1e-i)。茎组织石蜡纵切片分析表明,Ci-miR156c 超量表达植株的细胞直径显著小于对照,且单位横截面积内的细胞数量增加(图1j-l)。此外,Ci-miR156c 超量表达柑橘的开花时间也较对照组延迟。
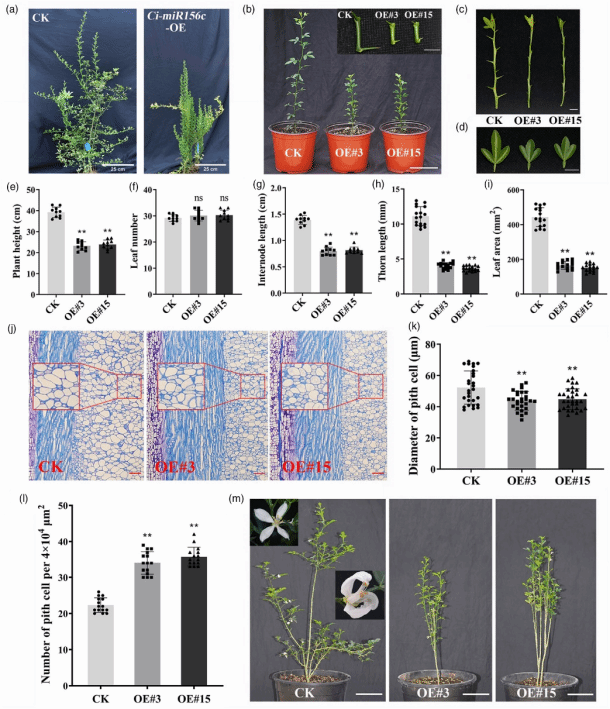
图1. Ci-miR156c-OE(过表达)转基因柑橘表型分析
2. Ci-miR156c通过靶向CiFD调控柑橘开花
前期研究已确定,Ci-miR156 在柑橘中靶向 9 个具有完整 SBP 结构域的 CiSPL 基因(Wu et al., 2010, 2015)。本研究通过 RT-qPCR 检测发现,在 Ci-miR156c 超量表达植株中,关键成花基因 CiFT、CiLFY、CiAP1 和 CiFD 的表达水平均较对照显著下调(图2a-d)。鉴于已有研究表明过表达 CiFT 和 CiFD 能促进柑橘开花(Wu et al., 2022; Ye et al., 2023),结合酵母单杂交(Y1H)、电泳迁移率变动分析(EMSA)及双荧光素酶报告基因(LUC)实验结果,本研究证实 Ci-miR156c-CiSPL7 模块通过靶向 CiFD 参与调控柑橘开花。
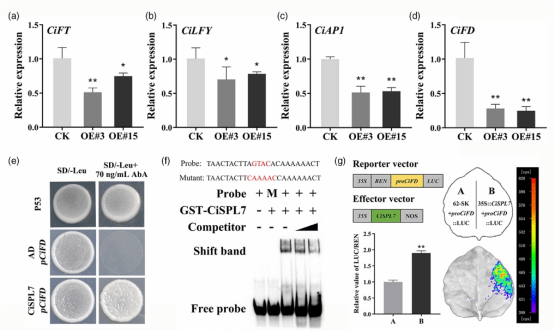
图2. Ci-miR156c-CiSPL模块通过靶向CiFD调控柑橘开花
3. CiKN6参与调控株高
研究发现 Ci-miR156c-CiSPL3 模块可能通过靶向 前期报道柑橘叶型发育关键调控基因CiKN6 参与株高发育调控(Zeng er al., 2022)。对 CiKN6 的功能分析显示,与对照相比,CiKN6 超量表达株系的株高、叶片数和平均节间长均显著降低(图3a-d)。茎组织石蜡纵切片分析表明,CiKN6 超量表达植株的细胞直径显著小于对照,且单位横截面积内的细胞数量增加(图3e-g)。利用病毒诱导基因沉默(VIGS)技术抑制柑橘中 CiKN6 的表达后,CiKN6-VIGS 植株的株高显著高于对照,而叶片数和平均节间长与对照无显著差异(图3h-k)。这些结果共同表明 CiKN6 参与调控柑橘株高。
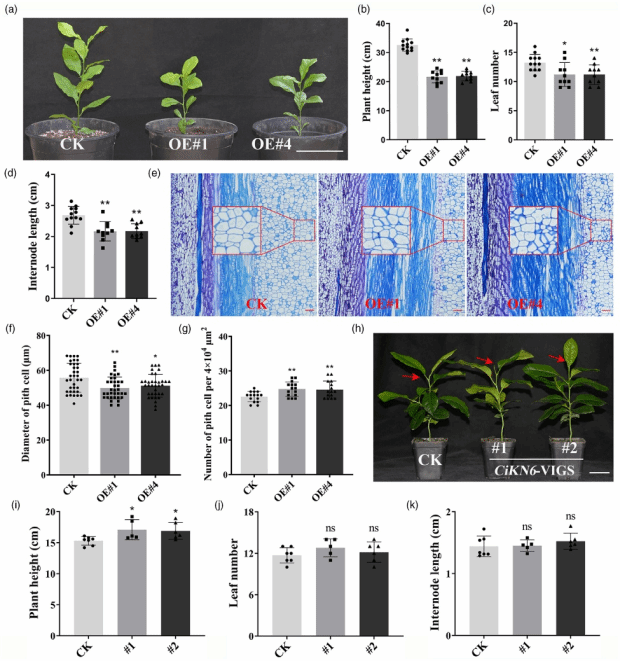
图3. CiKN6-OE(过表达)转基因柑橘的表型分析
4. Ci-miR156c和CiKN6表达改变了内源GA的含量
赤霉素(GA)通过促进细胞分裂和伸长调节株高,其在柑橘株高调控中的关键作用已被证实(Fagoaga et al., 2007)。RNA-seq 结果显示,Ci-miR156c 转基因植株与对照间的差异表达基因在激素水平调控、GA 应答以及 GA 介导的信号通路中显著富集。为探究 Ci-miR156c 和 CiKN6 转基因植株的矮化表型是否与 GA 含量变化相关,本研究测定了对照和转基因植株的内源 GA 含量。结果显示,在 Ci-miR156c-OE 转基因柑橘中,GA3 含量显著低于对照;而在 Ci-miR156c-STTM(Short Tandem Target Mimic)转基因柑橘中,GA3 含量则显著升高(图4a)。此外,CiKN6-OE 植株中的 GA3 含量也显著低于对照,而 CiKN6-VIGS 植株中的 GA3 含量略有升高(图4b, c)。值得注意的是,在这些转基因植株中均未检测到 GA1、GA4 和 GA7。这些结果表明,Ci-miR156c 和 CiKN6 表达的改变会影响内源 GA3 含量。
为分析 GA3 在株高调节中的作用,本研究分别用外源 GA3 和赤霉素抑制剂多效唑(paclobutrazol, PAC)处理转基因植株。GA3 处理使 Ci-miR156c-OE 株系的株高、叶片数和节间长度恢复至野生型(WT)水平(图4d-g)。与未经处理的 STTM 植株相比,PAC 处理显著降低了 Ci-miR156c-STTM 植株的株高(图4h-i),并减少了其叶片数量和平均节间长度(图4j-k)。同样地,GA3 处理也使 CiKN6-OE 株系的株高、叶片数和节间长度恢复至 WT 水平(图4l-o)。与之相反,PAC 处理显著降低了 CiKN6-VIGS 植株的株高(图4p-q),并减少了其叶片数量和平均节间长度(图4r-s)。综上所述,GA3 处理能完全恢复 Ci-miR156c-OE 和 CiKN6-OE 植株的矮化表型,表明这些转基因植株的矮化表型源于内源 GA3 水平的降低。
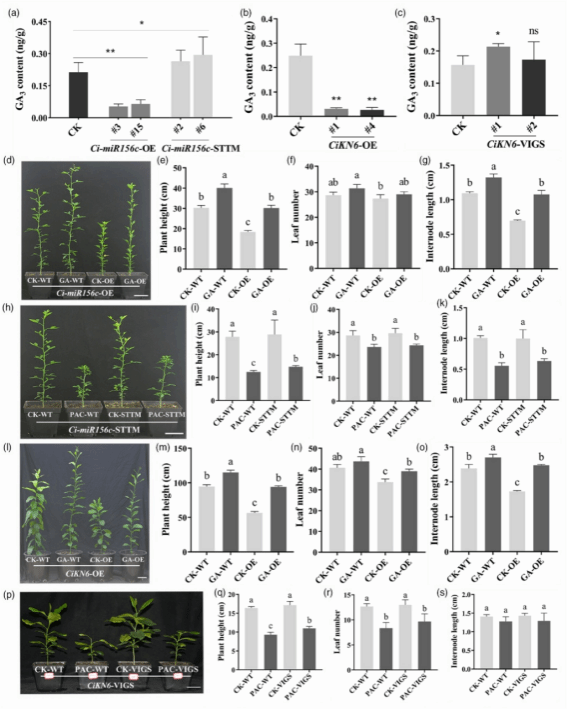
图4. GA3和PAC处理转基因植物的表型分析
5. Ci-miR156c-CiSPL 途径通过结合 CiGA20ox2 启动子抑制 GA 生物合成
已有研究证实,过表达 CiGA20ox2 可增加柑橘株高(Fagoaga et al., 2007; Kotoda et al., 2016; Vidal et al., 2001)。本研究的 RT-qPCR 结果显示,与对照相比,CiGA20ox2 在 Ci-miR156c-OE 和 CiKN6-OE 植株中表达下调,而在 Ci-miR156c-STTM 和 CiKN6-VIGS 植株中表达上调(图5a-b)。结合转基因植株中 GA3 含量和 CiGA20ox2 表达水平的变化,本研究推测 Ci-miR156c-CiSPL 模块可能直接或间接通过 Ci-miR156c-CiSPL3-CiKN6 途径调控 CiGA20ox2。进一步的 Y1H、EMSA、染色质免疫沉淀-qPCR(ChIP-qPCR)及 LUC 实验结果表明,Ci-miR156c-CiSPL6 途径可能直接结合 CiGA20ox2 启动子,或通过 Ci-miR156c-CiSPL3-CiKN6 途径间接调控其表达(图5c-h),从而抑制 Ci-miR156c-OE 和 CiKN6-OE 转基因植株中的 GA 生物合成。
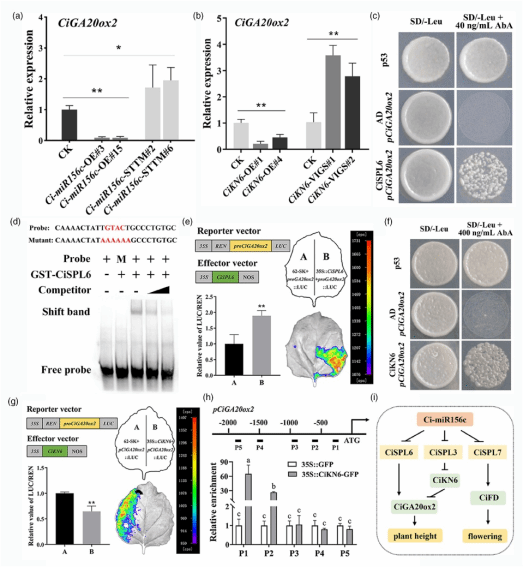
图5. Ci-miR156c-CiSPL路径通过结合CiGA20ox2启动子抑制GA的生物合成
基于以上结果,本研究提出了Ci-miR156c-CiSPL模块调控柑橘开花和株高的分子路径(图5i)。在开花调控中,Ci-miR156c靶向并抑制CiSPL7的表达,阻止其结合并激活CiFD的表达,从而延迟开花。在株高调控中,Ci-miR156c靶向并抑制CiSPL3的表达,CiSPL3与CiKN6启动子结合并抑制其表达。随后,CiKN6通过结合其自身启动子抑制GA生物合成基因CiGA20ox2的表达,最终减少植株内GA的生物合成。此外,Ci-miR156c抑制CiSPL6的表达,阻止其结合并激活CiGA20ox2的表达,进而导致植株内GA含量减少,从而抑制植株伸长。
华中农业大学张金智教授为该论文的通讯作者,胡春根教授对实验进行了全程指导。在读博士研究生谌敏为该论文的第一作者,已毕业博士张天亮、在读博士温永振、已毕业硕士张文博、马忠祥和席振萍参与课题工作。本研究得到了国家重点研发计划、国家自然科学基金和果蔬园艺作物种质创新与利用全国重点实验室的资助。
论文链接:https://doi.org/10.1111/pbi.70238
本文转自公众号:植物生物技术Pbj(PBJ | 华中农大破解柑橘“成花”与“矮化”协同调控密码,miR156-SPLs模块成关键靶点)




